



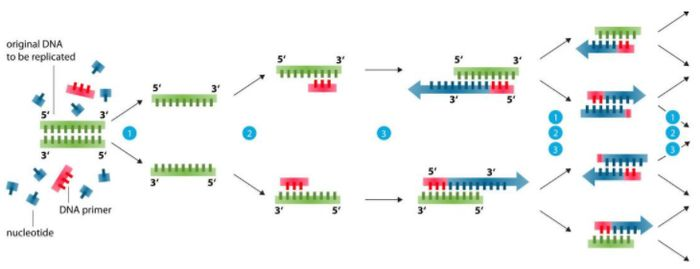
Krundi disaini alus (99% probleeme saab lahendada)
1. Aabitsa pikkus: õpik nõuab 15-30 bp, tavaliselt umbes 20 bp.Tegelik seisukord on spetsiifilisuse tagamiseks parem olla 18-24 bp, kuid mida pikem, seda parem, liiga pikk praimer vähendab ka spetsiifilisust ja vähendab saagist.
2. Praimeri amplifikatsiooni ulatus: 200-500 bp on sobiv ja fragmenti saab teatud tingimustel laiendada 10 kb-ni.
3. Praimeri alus: G+C sisaldus peaks olema 40-60%, liiga väike G+C võimendusefekt ei ole hea, liiga palju G+C on kergesti tekkivad mittespetsiifilised ribad.ATGC jaotub kõige paremini juhuslikult, vältides rohkem kui 5 puriini või pürimidiini nukleotiidi klastreid.Multi-gc 5'-otsa ja vahepealsete järjestuste jaoks, et suurendada stabiilsust, vältida rikkalikku GC-d 3'-otsas, ilma GC-d viimase 3 aluse jaoks või ilma GC-d 3-st viimasest 5-st alusest.
4. Vältige sekundaarset struktuuri praimerites ja vältige komplementeerimist kahe praimeri vahel, eriti komplementatsiooni 3' otsas, vastasel juhul moodustub praimeri dimeer ja tekivad mittespetsiifilised amplifitseeritud ribad.
5. Praimerite 3' otsas olevad alused, eriti viimased ja eelviimased alused, tuleks rangelt siduda, et vältida PCR-i ebaõnnestumist paaritute terminali aluste tõttu.
6. Praimeritel on või neid saab lisada sobivate lõhustamiskohtadega ja amplifitseeritud sihtjärjestusel peaksid eelistatavalt olema sobivad lõikamissaidid, mis on väga kasulik lõhustamisanalüüsi või molekulaarse kloonimise jaoks.
7. Praimerite spetsiifilisus: praimeritel ei tohiks olla ilmset homoloogiat teiste nukleiinhappejärjestuse andmebaasi järjestustega.
8. Õppige kasutama tarkvara: PP5, Oligo6, DNAstar, Vector NTI, primer3 (see veebikujundus töötab kõige paremini).
Ülaltoodud sisu võib lahendada vähemalt 99% praimeri disaini probleemidest.
Kontrollige krundi disaini üksikasju
1. Krundi pikkus
Üldine praimeri pikkus on 18-30 alust.Üldiselt on kõige olulisem praimeri lõõmutamistemperatuuri määrav tegur praimeri pikkus.Tavaliselt valitakse praimeri lõõmutamistemperatuur (Tm väärtus -5 ℃) ja mõned kasutavad otseselt Tm väärtust.Praimerite anniilimistemperatuuri ligikaudseks arvutamiseks saab kasutada järgmisi valemeid.
Kui praimeri pikkus on alla 20 bp: [4(G+C)+2(A+T)]-5 ℃
Kui praimeri pikkus on suurem kui 20 bp: 62,3 ℃ + 0,41 ℃ (%GC) -500/pikkus -5 ℃
Lisaks saab lõõmutamistemperatuuri arvutamiseks kasutada ka paljusid tarkvarasid, arvutuspõhimõte on erinev, nii et mõnikord võib arvutatud väärtuses olla väike vahe.PCR-reaktsioonide optimeerimiseks kasutatakse parima tõhususe ja spetsiifilisuse tagamiseks lühimaid praimereid, mis tagavad anniilimistemperatuuri vähemalt 54 ℃.
Üldiselt suureneb praimeri spetsiifilisus iga täiendava nukleotiidi puhul neli korda, nii et enamiku rakenduste minimaalne praimeri pikkus on 18 nukleotiidi.Praimeri pikkuse ülempiir ei ole eriti oluline, peamiselt seotud reaktsiooni efektiivsusega.Entroopia tõttu, mida pikem on praimer, seda madalam on selle anniilimise kiirus sihtmärk-DNA-ga seondumiseks, et moodustada stabiilne kaheahelaline matriitsi DNA polümeraasi seondumiseks.
Kui kasutate praimerite kujundamiseks tarkvara, saab praimerite pikkuse määrata omakorda TM väärtusega, eriti fluorestsents-kvantitatiivse PCR praimerite puhul tuleks kontrollida TM = 60 ℃ või nii.
2.GC sisu
Üldiselt on praimerite järjestustes G+C sisaldus 40% ~ 60% ning praimerite paari GC sisaldus ja Tm väärtus peaksid olema kooskõlastatud.Kui praimeril on tõsine GC või AT kalduvus, võib praimeri 5' otsa lisada sobiva koguse A, T või G ja C saba.
3. Lõõmutustemperatuur
Lõõmutamise temperatuur peaks olema 5 ℃ madalam kui lahtiahela temperatuur.Kui praimeri aluste arv on väike, saab anniilimistemperatuuri vastavalt tõsta, mis võib suurendada PCR spetsiifilisust.Kui aluste arv on suur, saab lõõmutamistemperatuuri vastavalt alandada.Anniilimistemperatuuri erinevus praimerite paari vahel 4 ℃ ~ 6 ℃ ei mõjuta PCR saagist, kuid ideaaljuhul on praimerite paari anniilimistemperatuur sama, mis võib varieeruda vahemikus 55 ℃ ~ 75 ℃.
4. Vältige võimendusmalli sekundaarse struktuuri ala
Amplifitseeritud fragmendi valimisel on kõige parem vältida malli sekundaarse struktuuri piirkonda.Sihtfragmendi stabiilset sekundaarset struktuuri saab ennustada ja hinnata asjakohase arvutitarkvara abil, mis on abiks malli valimisel.Katsetulemused näitavad, et paisumine on sageli ebaõnnestunud, kui paisutava piirkonna vaba energia (△G) on alla 58,6 lkJ/mol.
5. Mittevastavus sihtmärk-DNA-ga
Kui amplifitseeritud sihtmärk-DNA järjestus on suur, võib praimer seonduda sihtmärk-DNA mitme osaga, mille tulemuseks on mitu riba.Seekord on vaja kasutada BLAST tarkvara testimist, veebisaiti:http://www.ncbi.nlm.nih.gov/BLAST/.Valige Joonda kaks jada (bl2seq).
Praimeri järjestuste kleepimine tsooni 1 ja siht-DNA järjestuste kleepimine tsooni 2 on omavahel asendatav ning BLAST arvutab komplementaarsed, antisenss- ja muud võimalused, nii et kasutajad ei pea märkama, kas mõlemad ahelad on senssahelad.Kui teate andmebaasis oleva jada GI-numbrit, saate sisestada ka GI-numbri, nii et te ei pea kleepima suurt osa järjestusest.Lõpuks klõpsake Align at 3, et näha, kas praimeril on siht-DNA-s mitu homoloogset saiti.
6. Krundi terminal
Praimeri 3 'ots on koht, kus pikendus algab, seega on oluline vältida ebakõlade tekkimist.3' ots ei tohiks olla rohkem kui 3 järjestikust G või C, kuna see põhjustab praimeri ekslikult käivitamise G+C rikastusjärjestuse piirkonnas.3' ots ei saa moodustada sekundaarset struktuuri, välja arvatud spetsiaalsete PCR (AS-PCR) reaktsioonide korral, ei saa praimeri 3' ots olla sobimatu.Näiteks kui kodeerivat piirkonda amplifitseeritakse, ei tohiks praimeri 3' otsa lõpetada koodoni kolmandas positsioonis, kuna koodoni kolmas positsioon on kalduvus degenereeruda, mis mõjutab amplifikatsiooni spetsiifilisust ja efektiivsust.Anneksioonipraimerite kasutamisel vaadake koodonite kasutamise tabelit, pöörake tähelepanu bioloogilisele eelistusele, ärge kasutage anneksioonipraimereid 3′-otsas ja kasutage suuremat kontsentratsiooni praimereid (1 uM-3 uM).
7. Praimerite sekundaarne struktuur
Praimeritel endil ei tohiks olla komplementaarseid järjestusi, vastasel juhul voldivad praimerid ise juuksenõelastruktuurideks ja see sekundaarne struktuur mõjutab praimerite ja mallide seondumist steerilise takistuse tõttu.Kui kasutatakse kunstlikku otsust, ei tohiks praimerite endi pidevad komplementaarsed alused olla suuremad kui 3 bp.Kahe praimeri vahel ei tohiks olla komplementaarsust, eriti tuleks vältida 3' otsa komplementaarset kattumist, et vältida praimeri dimeeride moodustumist.Üldiselt ei tohiks praimerite paari vahel olla rohkem kui 4 järjestikust aluse homoloogiat või komplementaarsust.
8. Lisa markerid või lookused
5'-otsal on amplifikatsioonispetsiifilisusele väike mõju ja seetõttu saab seda modifitseerida ilma amplifikatsioonispetsiifilisust mõjutamata.Praimeri 5' otsa modifikatsioon hõlmas: ensüümi restriktsioonisaidi lisamist;Märgistatud biotiin, fluorestsents, digoksiin, Eu3+ jne. Sisestage valku siduvad DNA järjestused;Mutatsioonikohtade sisseviimine, mutatsioonijärjestuste sisestamine ja puudumine ning promootorjärjestuste sisseviimine jne. Lisaalused mõjutavad enam-vähem amplifikatsiooni efektiivsust ja suurendavad praimeri dimeeri moodustumise võimalust, kuid järgmise sammu osas tuleb teha mõningaid mööndusi.Täiendavaid järjestusi, mida sihtjärjestuses ei eksisteeri, nagu restriktsioonisaidid ja promootorjärjestused, saab lisada praimeri 5'-otsa ilma spetsiifilisust mõjutamata.Neid järjestusi ei kaasata praimeri Tm väärtuste arvutamisse, kuid neid tuleks testida komplementaarsuse ja sisemise sekundaarstruktuuri suhtes.
9. Alamkloonid
Enamasti on PCR ainult esialgne kloonimine ja seejärel peame sihtfragmendi subkloonima erinevatesse vektoritesse, nii et peame kavandama täiendavad alused PCR-i etapi järgmiseks operatsiooniks.
Mõned subkloonimiseks mõeldud järjestused on kokku võetud allpool.
Restriktsiooni endonukleaasi restriktsioonisait lisati
Ensüümi restriktsioonisaitide lisamine on kõige sagedamini kasutatav meetod PCR-produktide subkloonimiseks.Üldiselt on lõhustamiskohaks kuus alust, lisaks lõhustamiskoha 5 'otsale tuleb lisada 2–3 kaitsvat alust.Erinevate ensüümide jaoks vajalike kaitsvate aluste arv on aga erinev.Näiteks SalⅠ ei vaja kaitsealust, EcoRⅤ vajab 1 kaitsealust, NotⅠ vajab 2 kaitsealust ja Hind Ⅲ vajab 3 kaitsealust.
LIC lisab saba
LIC täisnimi on Ligation-Independent kloonimine, Navogeni poolt leiutatud kloonimismeetod spetsiaalselt selle osa jaoks pET-vektorist.LIC-meetodil valmistatud pET-kandjal on mittekomplementaarsed 12-15-aluselised üheahelalised kleepuvad otsad, mis täiendavad sihtmärk-inserdi fragmendi vastavaid kleepuvaid otsi.Amplifikatsiooni eesmärgil peaks sisestatud fragmendi praimeri 5' järjestus täiendama LIC-vektorit.T4 DNA polümeraasi 3′→5′ ekstranektne aktiivsus võib lühikese aja pärast moodustada sisestatud fragmendile üheahelalise kleepuva otsa.Kuna produkti saab moodustada ainult ettevalmistatud inserdi fragmendi ja vektori vastastikusel anniilimisel, on see meetod väga kiire ja tõhus ning see on suunatud kloonimine.
Suunatud TA kloon lisa saba
TA kloonimine ei suutnud fragmenti vektorisse suunata, nii et hiljem tutvustas Invitrogen vektori, mis võis sihtida kloonimist, mis sisaldas ühes otsas nelja silmapaistvat GTGGS-i.Seetõttu tuleks PCR-praimerite kavandamisel vastavalt lisada komplementaarsed järjestused, et fragmente saaks "orienteerida".
Kui teil napib aega, võite proovida otsest sünteesi, kombineerides geeni vektoriga, mida me nimetame musekularistides ET geeni sünteesiks.
D. In-Fusion kloonimise meetod
Pole vaja ligaasi, pole vaja pikka reaktsiooni.Seni, kuni sisestatakse järjestus lineariseeritud vektori mõlemas otsas Praimerite konstrueerimisel lisatakse seejärel PCR produkt ja lineariseeritud vektor BSA-d sisaldavasse infusiooniensüümi lahusesse ja asetatakse pooleks tunniks toatemperatuurile, saab transformatsiooni läbi viia.See meetod sobib eriti hästi suure mahu muundamiseks.
10. Ühendage praimer
Mõnikord on praimeri disaini kohta teada ainult piiratud järjestusteave.Näiteks kui on teada ainult aminohappejärjestus, saab kavandada ühineva praimeri.Ühinemispraimer on segu erinevatest järjestustest, mis esindavad kõiki erinevaid aluseid, mis kodeerivad ühte aminohapet.Spetsiifilisuse suurendamiseks võite viidata koodonikasutuse tabelile, et vähendada anneksiooni vastavalt erinevate organismide baaskasutuse eelistustele.Hüpoksantiini saab siduda kõigi alustega, et vähendada praimeri anniilimistemperatuuri.Ärge kasutage lisatud aluseid praimeri 3′ otsas, sest viimase 3 aluse anniilimine 3′ otsas on piisav, et käivitada PCR vales kohas.Kasutatakse kõrgemaid praimerite kontsentratsioone (1 μM kuni 3 μM), kuna paljude anneksioonisegude praimerid ei ole sihtmatriitsile spetsiifilised.
PCR toorainedkontroll
1. Krundi kogus
Iga praimeri kontsentratsioon on 0,1-1 umol või 10-100 pmol.Parem on saada vajalik tulemus väikseima kruntvärviga.Praimeri kõrge kontsentratsioon põhjustab mittevastavust ja mittespetsiifilist amplifikatsiooni ning suurendab praimerite vahel dimeeride moodustumise võimalust.
2. Praimeri kontsentratsioon
Praimerite kontsentratsioon mõjutab spetsiifilisust.Optimaalne praimeri kontsentratsioon on üldiselt vahemikus 0,1 kuni 0,5 μM.Kõrgemad praimerite kontsentratsioonid põhjustavad mittespetsiifiliste produktide amplifikatsiooni.
3. Krundi lõõmutamistemperatuur
Teine oluline praimerite parameeter on sulamistemperatuur (Tm).See on temperatuur, kui 50% praimeritest ja komplementaarsetest järjestustest on esindatud kaheahelaliste DNA molekulidena.PCR anniilimistemperatuuri määramiseks on vajalik Tm.Ideaalis on anniilimistemperatuur piisavalt madal, et tagada praimerite tõhus anniilimine sihtjärjestusega, kuid piisavalt kõrge, et vähendada mittespetsiifilist seondumist.Mõistlik lõõmutamistemperatuur vahemikus 55 ℃ kuni 70 ℃.Lõõmutamistemperatuur on üldiselt seatud 5 ℃ madalamaks kui krundi Tm.
Tm seadistamiseks on mitu valemit, mis olenevalt kasutatavast valemist ja praimerite järjestusest erinevad suuresti.Kuna enamik valemeid annab hinnangulise Tm väärtuse, on kõik lõõmutamistemperatuurid ainult lähtepunktiks.Spetsiifilisust saab parandada, analüüsides mitmeid reaktsioone, mis järk-järgult tõstavad anniilimistemperatuuri.Alustage alla hinnangulise Tm-5 ℃ ja tõstke lõõmutamistemperatuuri järk-järgult 2 ℃ võrra.Kõrgem anniilimistemperatuur vähendab praimerite dimeeride ja mittespetsiifiliste toodete teket.Parimate tulemuste saavutamiseks peaksid kahe praimeri Tm väärtused olema ligikaudsed.Kui praimeripaaride Tm erinevus on suurem kui 5 ℃, näitavad praimerid olulist valekäivitust, kasutades tsüklis madalamat anniilimistemperatuuri.Kui kaks praimerit Tm on erinevad, seadke anniilimistemperatuur 5 ℃ madalamale kui madalaim Tm.Alternatiivina võib spetsiifilisuse suurendamiseks läbi viia viis tsüklit kõigepealt kõrgema Tm jaoks mõeldud lõõmutamistemperatuuridel, millele järgneb ülejäänud tsüklid madalama Tm jaoks mõeldud lõõmutamistemperatuuridel.See võimaldab kitsastes tingimustes saada sihtmalli osalise koopia.
4. Praimeri puhtus ja stabiilsus
Kohandatud praimerite standardne puhtus on enamiku PCR rakenduste jaoks piisav.Bensoüül- ja isobutüülrühmade eemaldamine soola eemaldamise teel on minimaalne ja seetõttu ei sega see PCR-i.Mõned rakendused nõuavad puhastamist, et eemaldada sünteesiprotsessis kõik mittetäispikad järjestused.Need kärbitud järjestused tekivad seetõttu, et DNA sünteesi keemia efektiivsus ei ole 100%.See on ümmargune protsess, mis kasutab korduvaid keemilisi reaktsioone, kuna iga alus lisatakse DNA muutmiseks 3′ kuni 5′.Mõlemas tsüklis võite ebaõnnestuda.Pikematel praimeritel, eriti neil, mis on suuremad kui 50 alust, on suur osa kärbitud järjestusi ja need võivad vajada puhastamist.
Praimerite saagist mõjutavad sünteetilise keemia ja puhastusmeetodi efektiivsus.Biofarmatseutilised ettevõtted, nagu Cytology ja Shengong, kasutavad oligonukleosiidide kogutoodangu tagamiseks minimaalset OD-ühikut.Kohandatud praimerid tarnitakse kuivpulbri kujul.Kõige parem on praimerid uuesti lahustada TE-s nii, et lõppkontsentratsioon oleks 100 μM.TE on parem kui deioniseeritud vesi, kuna vee pH on sageli happeline ja põhjustab oligonukleosiidide hüdrolüüsi.
Kruntvärvide stabiilsus sõltub säilitustingimustest.Kuivpulbrit ja lahustunud praimereid tuleb hoida temperatuuril -20 ℃.TE-s lahustatud praimereid, mille kontsentratsioon on suurem kui 10 μM, saab stabiilselt säilitada temperatuuril -20 ℃ 6 kuud, kuid toatemperatuuril (15 ℃ kuni 30 ℃) võib neid hoida ainult vähem kui 1 nädal.Kuivpulbri kruntvärvid säilivad -20 C juures vähemalt 1 aasta ja toatemperatuuril (15 C kuni 30 C) kuni 2 kuud.
5. Ensüümid ja nende kontsentratsioonid
Praegu on kasutatav Taq DNA polümeraas põhiliselt kolibakterite poolt sünteesitud geenitehnoloogia ensüüm.Tüüpilise PCR-reaktsiooni katalüüsimiseks vajalik ensüümi kogus on umbes 2,5 U (viitab reaktsiooni kogumahule 100 ul).Kui kontsentratsioon on liiga kõrge, võib see põhjustada mittespetsiifilist võimendust;kui kontsentratsioon on liiga madal, vähendatakse sünteetilise toote kogust.
6. dNTP kvaliteet ja kontsentratsioon
dNTP kvaliteet on tihedalt seotud PCR amplifikatsiooni kontsentratsiooni ja efektiivsusega.dNTP pulber on granuleeritud ja selle varieeruvus kaotab oma bioloogilise aktiivsuse, kui seda valesti hoitakse.dNTP lahus on happeline ja seda tuleks kasutada kõrge kontsentratsiooniga koos 1 M NaOH või 1 M Tris.HCL puhverlahusega, et reguleerida selle pH väärtusele 7,0–7,5, väike kogus alampakendit, külmutatud säilitamine temperatuuril -20 ℃.Mitmekordne külmutamine-sulatamine halvendab dNTP-d.PCR reaktsioonis peaks dNTP olema 50–200 umol/l.Eriti tuleks tähelepanu pöörata sellele, et nelja DNTPS-i kontsentratsioon peaks olema võrdne (võrdne moolipreparaat).Kui mõne neist kontsentratsioon erineb teistest (kõrgem või madalam), tekib mittevastavus.Liiga madal kontsentratsioon vähendab PCR-produktide saagist.dNTP võib kombineerida Mg2+-ga ja vähendada vaba Mg2+ kontsentratsiooni.
7. Matriits (sihtgeen) nukleiinhape
Matriitsi nukleiinhappe kogus ja puhastusaste on PCR õnnestumise või ebaõnnestumise üks peamisi lülisid.Traditsioonilised DNA puhastusmeetodid kasutavad proovide seedimiseks ja kõrvaldamiseks tavaliselt SDS-i ja proteaasi K.SDS-i põhifunktsioonid on: lahustada rakumembraanil olevaid lipiide ja valke, hävitades seeläbi rakumembraani lahustades membraanivalke, ning dissotsieerida rakus tuumavalke, SDS võib ka valkudega kombineerida ja sadestada;Proteaas K võib hüdrolüüsida ja seedida valke, eriti DNA-ga seotud histoone ning seejärel kasutada valkude ja muude rakukomponentide ekstraheerimiseks orgaanilist lahustit fenooli ja kloroformi ning nukleiinhappe sadestamiseks kasutada etanooli või isopropüülalkoholi.Ekstraheeritud nukleiinhapet saab kasutada PCR reaktsioonide matriitsina.Üldise kliinilise tuvastamise proovide puhul saab kiiret ja lihtsat meetodit kasutada rakkude lahustamiseks, patogeenide lüseerimiseks, kromosoomidest valkude seedimiseks ja eemaldamiseks vabade sihtgeenideni ning seda kasutatakse otse PCR amplifikatsiooniks.RNA matriitsi ekstraheerimiseks kasutatakse tavaliselt guanidiini isotiotsüanaadi või proteaasi K meetodit, et vältida RNAasi RNA lagundamist.
8.Mg2+ kontsentratsioon
Mg2+ mõjutab oluliselt PCR amplifikatsiooni spetsiifilisust ja saagist.Üldises PCR reaktsioonis, kui erinevate dNTP kontsentratsioon on 200 umol/l, on sobiv Mg2+ kontsentratsioon 1,5 ~ 2,0 mmol/l.Mg2+ kontsentratsioon on liiga kõrge, reaktsiooni spetsiifilisus väheneb, toimub mittespetsiifiline amplifikatsioon, liiga madal kontsentratsioon vähendab Taq DNA polümeraasi aktiivsust, mille tulemuseks on reaktsiooniproduktide vähenemine.
Magneesiumioonid mõjutavad PCR-i mitmeid aspekte, nagu DNA polümeraasi aktiivsus, mis mõjutab saagist;Teine näide on praimeri anniilimine, mis mõjutab spetsiifilisust.dNTP ja matriits seonduvad magneesiumiooniga, vähendades ensüümi aktiivsuseks vajaliku vaba magneesiumiooni hulka.Optimaalne magneesiumioonide kontsentratsioon erineb erinevate praimeripaaride ja mallide puhul, kuid tüüpiline PCR algkontsentratsioon 200 μM dNTP-ga on 1,5 mM (märkus: reaalajas kvantitatiivse PCR jaoks kasutage fluorestsentssondiga 3–5 mM magneesiumioonide lahust).Vabade magneesiumiioonide kõrgem kontsentratsioon suurendab saagist, kuid suurendab ka mittespetsiifilist amplifikatsiooni ja vähendab täpsust.Optimaalse kontsentratsiooni määramiseks viidi magneesiumiioonide tiitrimine läbi 0,5 mM sammuga 1 mM kuni 3 mM.Magneesiumioonide optimeerimisest sõltuvuse vähendamiseks võib kasutada Platinum Taq DNA polümeraasi.Platinum Taq DNA polümeraas suudab säilitada funktsiooni laiemas magneesiumioonide kontsentratsioonide vahemikus kui Taq DNA polümeraas ja vajab seetõttu vähem optimeerimist.
9. Pcr-i soodustavad lisandid
Lõõmutamistemperatuuri, praimeri disaini ja magneesiumiioonide kontsentratsiooni optimeerimisest piisab enamiku mallide väga spetsiifiliseks võimendamiseks;mõned mallid, sealhulgas suure GC-sisaldusega mallid, nõuavad siiski lisameetmeid.Lisandid, mis mõjutavad DNA sulamistemperatuuri, on veel üks viis toote spetsiifilisuse ja saagise parandamiseks.Parimate tulemuste saavutamiseks on vajalik malli täielik denatureerimine.
Lisaks takistab sekundaarne struktuur praimeri sidumist ja ensüümi pikendamist.
PCR lisandid, sealhulgas formamiid, DMSO, glütseriin, betaiin ja PCRx Enhancer Solution, suurendavad võimendamist.Nende võimalikuks mehhanismiks on sulamistemperatuuri alandamine, aidates seega kaasa praimerite anniilimisele ja abistades DNA polümeraasi pikendamist läbi sekundaarstruktuuri piirkonna.PCRx lahendusel on muid eeliseid.Plaatinum Taq DNA polümeraasi ja Platinum Pfx DNA polümeraasi kasutamisel on vajalik minimaalne magneesiumioonide optimeerimine.Seega kombineeritakse Platinum tehnikat lisandiga, et suurendada spetsiifilisust, vähendades samas sõltuvust kolmandast lähenemisviisist, magneesiumioonide optimeerimisest.Parimate tulemuste saavutamiseks tuleks optimeerida lisandite, eriti DMSO, formamiidi ja glütserooli kontsentratsiooni, mis inhibeerivad Taq DNA polümeraasi.

10. Kuum algus
Kuumkäivituse PCR on lisaks heale praimeri disainile üks olulisemaid meetodeid PCR spetsiifilisuse parandamiseks.Kuigi Taq DNA polümeraasi optimaalne pikenemistemperatuur on 72 ℃, jääb polümeraas aktiivseks ka toatemperatuuril.Seega tekivad mittespetsiifilised produktid, kui säilitustemperatuur on PCR reaktsiooni valmistamise ajal ja termilise tsükli alguses anniilimistemperatuurist madalam.Pärast moodustumist võimenduvad need mittespetsiifilised tooted tõhusalt.Kuumkäivitusega PCR on eriti tõhus, kui praimeri kujundamiseks kasutatavad saidid on piiratud geneetiliste elementide asukohaga, nagu saidipõhised mutatsioonid, ekspressioonikloonimine või DNA inseneriks kasutatavate geneetiliste elementide konstrueerimine ja manipuleerimine.
Tavaline meetod Taq DNA polümeraasi aktiivsuse piiramiseks on valmistada jääl PCR reaktsioonilahus ja asetada see eelsoojendatud PCR aparaati.See meetod on lihtne ja odav, kuid see ei vii ensüümi aktiivsust lõpuni ega välista seetõttu täielikult mittespetsiifiliste toodete võimendamist.
Termiline praimimine aeglustab DNA sünteesi, inhibeerides olulist komponenti, kuni PCR-seade saavutab denaturatsioonitemperatuuri.Enamik käsitsi termilise initsiatsiooni meetodeid, sealhulgas Taq DNA polümeraasi viivitatud lisamist, on tülikad, eriti suure läbilaskevõimega rakenduste puhul.Teiste termiliste kruntimismeetodite puhul kasutatakse olulise komponendi, sealhulgas magneesiumioonide või ensüümide ümbritsemiseks või reaktiivsete komponentide, näiteks mallide ja puhvrite füüsiliseks isoleerimiseks vahakaitset.Termilise tsükli käigus vabanevad erinevad komponendid ja segunevad vaha sulamisel.Nagu manuaalne kuumkäivitusmeetod, on ka vahakaitsemeetod tülikas ja saastumisele kalduv ning ei sobi suure läbilaskevõimega rakenduste jaoks.
Plaatina DNA polümeraas on mugav ja tõhus automaatse kuumkäivitusega PCR-i jaoks.Platinum Taq DNA polümeraas koosneb rekombinantsest Taq DNA polümeraasist, mis on kombineeritud Taq DNA polümeraasi vastaste monoklonaalsete antikehadega.Antikehad formuleeritakse PCR abil, et inhibeerida ensüümi aktiivsust pikaajalisel temperatuuri hoidmisel.Taq DNA polümeraas vabanes reaktsioonise denaturatsioonietapi 94 ℃ isolatsiooni ajal, taastades täieliku polümeraasi aktiivsuse.Vastupidiselt termilise initsiatsiooniga keemiliselt modifitseeritud Taq DNA polümeraasile ei vaja plaatina ensüüm polümeraasi aktiveerimiseks pikemaajalist isolatsiooni temperatuuril 94 ℃ (10 kuni 15 minutit).PlatinumTaq DNA polümeraasiga taastati 90% Taq DNA polümeraasi aktiivsusest 2 minuti pärast temperatuuril 94 ℃.

11. Nest-PCR
Järjestikused amplifikatsioonivoorud, kasutades pesastatud praimereid, võivad parandada spetsiifilisust ja tundlikkust.Esimene ring on standardvõimendus 15-20 tsüklit.Väike osa esialgsest amplifikatsiooniproduktist lahjendati 100 kuni 1000 korda ja lisati 15 kuni 20 tsükliks teise amplifikatsioonivooru.Alternatiivselt võib esialgse amplifitseeritud produkti suuruse määrata geelpuhastusega.Teises amplifikatsioonivoorus kasutatakse pesastatud praimerit, mis võib seostuda esimese praimeri sees oleva sihtjärjestusega.Pesastatud PCR kasutamine vähendab mitme sihtmärgi amplifikatsiooni võimalust, kuna mõlema praimerite komplektiga komplementaarseid sihtjärjestusi on vähe.Sama tsüklite koguarv (30 kuni 40) samade praimeritega võimendas mittespetsiifilisi saite.Pesastatud PCR suurendab piiratud sihtmärkjärjestuste (nt haruldaste mrnas) tundlikkust ja parandab keerulise PCRS (nt 5' RACE) spetsiifilisust.
12. Langev PCR
Langev PCR parandab spetsiifilisust, kasutades PCR-i esimeste tsüklite jaoks tihedaid anniilimistingimusi.Tsükkel algab lõõmutamistemperatuuril, mis on ligikaudu 5 ℃ kõrgem kui hinnanguline Tm, seejärel vähendatakse iga tsüklit 1 ℃ kuni 2 ℃ võrra, kuni lõõmutamistemperatuur on alla Tm 5 ℃.Võimendatakse ainult suurima homoloogiaga sihtkoha malli.Need tooted jätkavad laienemist järgmiste tsüklite jooksul, tõrjudes välja võimendatud mittespetsiifilised tooted.Langev PCR on kasulik meetodite puhul, kus praimeri ja sihtmatriitsi homoloogia aste ei ole teada, näiteks AFLP DNA sõrmejälgede võtmine.
Seotud PCR komplektid
2× PCR kangelaneTMSegamissüsteemil on suurem tolerantsus PCR inhibiitorite suhtes kui tavalisel PCR Mix süsteemil ja see saab hõlpsasti hakkama erinevate komplekssete mallide PCR amplifikatsiooniga.Ainulaadne reaktsioonisüsteem ja kõrge efektiivsusega Taq Hero muudavad PCR-reaktsioonil kõrgema amplifikatsioonitõhususe, spetsiifilisuse ja tundlikkuse.
Kõrgem võimenduse efektiivsus
Sellel on 5'→3' DNA polümeraasi aktiivsus ja 5'→3' eksonukleaasi aktiivsus, ilma 3'→5' eksonukleaasi aktiivsuseta.
Reaalajas PCR Easyᵀᴹ-SYBR Green I komplekt
Spetsiifiline optimeeritud puhver ja kuumkäivitusega Taq ensüüm võivad takistada mittespetsiifilist amplifikatsiooni ja praimeri dimeeri moodustumist
Kõrge tundlikkus – suudab tuvastada madalaid mallikoopiaid
Komplektis kasutatakse ainulaadset Foregene pöördtranskriptsiooni reaktiivi ja Foregene HotStar Taq DNA polümeraasi kombineerituna ainulaadse reaktsioonisüsteemiga, et tõhusalt parandada amplifikatsiooni efektiivsust ja reaktsiooni spetsiifilisust.
Postitusaeg: mai-09-2023



