



一、Suurendage reaktsioonisüsteemi tundlikkust:
1. Eraldage kõrge kvaliteediga RNA:
Edukas cDNA süntees pärineb kvaliteetsest RNA-st.Kvaliteetne RNA peaks olema vähemalt täispikk ja vaba pöördtranskriptaasi inhibiitoritest nagu EDTA või SDS.RNA kvaliteet määrab järjestusteabe maksimaalse hulga, mida saate cDNA-sse transkribeerida.Tavaline RNA puhastamise meetod on üheetapiline meetod, milles kasutatakse guanidiini isotiotsüanaati/happefenooli.RNaasi jälgede saastumise vältimiseks tuleb RNaasi sisaldavatest proovidest (näiteks kõhunäärmest) eraldatud RNA-d säilitada formaldehüüdis, et säilitada kvaliteetne RNA, eriti pikaajaliseks säilitamiseks.Roti maksast ekstraheeritud RNA lagunes põhimõtteliselt pärast ühenädalast vees hoidmist, samas kui roti põrnast ekstraheeritud RNA püsis pärast 3-aastast vees hoidmist stabiilsena.Lisaks on üle 4 kb pikkused transkriptid tundlikumad RNaasi jälgede lagunemise suhtes kui väikesed transkriptid.Säilitatud RNA proovide stabiilsuse suurendamiseks võib RNA lahustada deioniseeritud formamiidis ja säilitada temperatuuril -70 °C.RNA säilitamiseks kasutatav formamiid peab olema vaba RNA-d lagundavast prahist.Pankrease RNA-d saab vormimiidis säilitada vähemalt ühe aasta.RNA kasutamiseks valmistudes saate RNA sadestamiseks kasutada järgmist meetodit: lisage NaCl 0,2 M ja 4-kordsele etanooli mahule, asetage 3–5 minutiks toatemperatuurile ja tsentrifuugige 10 000 × g juures 5 minutit.
2. Kasutage RNaseH-inaktiivset (RNaseH-) pöördtranskriptaasi:
Sageli lisatakse pöördtranskriptsioonireaktsioonidele RNaasi inhibiitoreid, et suurendada cDNA sünteesi pikkust ja saagist.RNaasi inhibiitorid tuleks lisada esimese ahela sünteesireaktsiooni ajal puhvri ja redutseeriva aine (nt DTT) juuresolekul, kuna cDNA sünteesile eelnev protsess denatureerib inhibiitori, vabastades sellega seotud RNaasi, mis võib RNA-d lagundada.Valgu RNaasi inhibiitorid takistavad ainult RNA lagunemist RNaasi A, B, C poolt ega takista RNaasi sattumist nahale, seega olge ettevaatlik, et vaatamata nende inhibiitorite kasutamisele ei satuks RNaasi oma sõrmedest.
Pöördtranskriptaas katalüüsib RNA konversiooni cDNA-ks.Nii M-MLV kui ka AMV omavad endogeenset RNaasH aktiivsust lisaks nende enda polümeraasi aktiivsusele.RNaasH aktiivsus ja polümeraasi aktiivsus konkureerivad üksteisega RNA matriitsi ja DNA praimeri või cDNA pikendusahela vahel moodustunud hübriidahela pärast ning lagundavad RNA ahelat RNA:DNA kompleksis.RNaasH aktiivsusega lagunenud RNA matriits ei saa enam olla cDNA sünteesi tõhusa substraadina, mis vähendab cDNA sünteesi saagist ja pikkust.Seetõttu oleks kasulik kõrvaldada või oluliselt vähendada pöördtranskriptaasi RNaasH aktiivsust.
SuperScript Ⅱ pöördtranskriptaas, RNaseH-MMLV pöördtranskriptaas ja thermoScript pöördtranskriptaas, RNaseH-AMV, suudavad saada rohkem kogust ja rohkem täispikka cDNA-d kui MMLV ja AMV.RT-PCR tundlikkust mõjutab cDNA sünteesi kogus.ThermoScript on palju tundlikum kui AMV.RT-PCR produktide suurust piirab pöördtranskriptaasi võime cDNA-d sünteesida, eriti suuremate cDNA-de kloonimisel.Võrreldes MMLV-ga suurendas SuperScripⅡ märkimisväärselt pikkade RT-PCR toodete saagist.RNaseH-pöördtranskriptaasil on ka suurem termostabiilsus, nii et reaktsiooni saab läbi viia normaalsest 37–42 °C kõrgematel temperatuuridel.Soovitatud sünteesitingimustes kasutage oligo(dT) praimerit ja 10 μCi [α-P]dCTP-d.Esimese ahela kogusaagis arvutati TCA sadestamise meetodil.Täispikka cDNA-d analüüsiti, kasutades suuruse järgi sorteeritud ribasid, mis lõigati välja ja loendati leeliselisel agaroosgeelil.
3. Tõstke pöördtranskriptsiooni inkubatsioonitemperatuuri:
Kõrgem inkubatsioonitemperatuur aitab avada RNA sekundaarset struktuuri, suurendades reaktsiooni saagist.Enamiku RNA matriitside puhul kõrvaldab RNA ja praimerite inkubeerimine 65 °C juures ilma puhvri või soolata, millele järgneb kiire jahutamine jääl enamiku sekundaarstruktuuridest ja võimaldab praimeritel seonduda.Mõnel mallil on siiski sekundaarsed struktuurid, isegi pärast kuumdenatureerimist.Neid keerulisi malle saab amplifitseerida, kasutades ThermoScripti pöördtranskriptaasi ja asetades amplifikatsiooni parandamiseks pöördtranskriptsioonireaktsiooni kõrgemale temperatuurile.Kõrgem inkubatsioonitemperatuur võib samuti suurendada spetsiifilisust, eriti kui cDNA sünteesiks kasutatakse geenispetsiifilisi praimereid (GSP) (vt 3. peatükk).Kui kasutate GSP-d, veenduge, et praimerite Tm on sama, mis eeldatav inkubatsioonitemperatuur.Ärge kasutage oligo(dT) ja juhuslikke praimereid temperatuuril üle 60 °C.Juhuslikud praimerid vajavad inkubeerimist 25 °C juures 10 minutit enne temperatuuri tõstmist 60 °C-ni.Lisaks kõrgema pöördtranskriptsiooni temperatuuri kasutamisele saab spetsiifilisust parandada ka RNA/praimeri segu otse ülekandmisega 65 °C denatureerimistemperatuurilt pöördtranskriptsiooni inkubatsioonitemperatuurile ja eelsoojendatud 2x reaktsioonisegu lisamisega (cDNA kuumkäivitussüntees).See lähenemisviis aitab vältida molekulidevahelist aluste paaristumist, mis toimub madalamatel temperatuuridel.RT-PCR jaoks vajalikku temperatuuri mitmekordset ümberlülitamist saab lihtsustada termotsükleri abil.
Tth termostabiilne polümeraas toimib Mg2+ juuresolekul DNA polümeraasina ja Mn2+ juuresolekul RNA polümeraasina.Seda saab hoida soojas kuni 65°C.Kuid Mn2+ olemasolu PCR-i ajal vähendab täpsust, mis muudab Tth polümeraasi vähem sobivaks ülitäpse amplifikatsiooni jaoks, näiteks cDNA kloonimiseks.Lisaks on Tth-l madal pöördtranskriptsiooni efektiivsus, mis vähendab tundlikkust, ja kuna pöördtranskriptsiooni ja PCR-i saab läbi viia ühe ensüümiga, ei saa pöördtranskriptsioonita kontrollreaktsioone kasutada cDNA amplifikatsiooniproduktide võrdlemiseks saastava genoomse DNA-ga.Amplifikatsiooniproduktid eraldati.
4. Lisandid, mis soodustavad pöördtranskriptsiooni:
Esimese ahela sünteesireaktsioonile lisatakse lisaaineid, sealhulgas glütserooli ja DMSO-d, mis võivad vähendada nukleiinhappe kaheahelalise stabiilsust ja lahti siduda RNA sekundaarse struktuuri.SuperScript II või MMLV aktiivsust mõjutamata võib lisada kuni 20% glütserooli või 10% DMSO-d.AMV talub ka kuni 20% glütserooli aktiivsust kaotamata.RT-PCR tundlikkuse maksimeerimiseks SuperScriptⅡ pöördtranskriptsiooni reaktsioonis võib lisada 10% glütserooli ja inkubeerida 45 °C juures.Kui PCR-ile lisatakse 1/10 pöördtranskriptsiooni reaktsiooni produktist, siis glütserooli kontsentratsioon amplifikatsioonireaktsioonis on 0,4%, millest ei piisa PCR inhibeerimiseks.
5. RNaasH ravi:
cDNA sünteesireaktsioonide töötlemine RNaasH-ga enne PCR-i võib suurendada tundlikkust.Mõnede mallide puhul arvatakse, et cDNA sünteesireaktsioonis olev RNA takistab amplifikatsiooniproduktide seondumist, sel juhul võib ravi RNaasH-ga suurendada tundlikkust.Üldiselt on ravi RNaasH-ga vajalik pikemate täispikkade cDNA sihtmärkide, näiteks väikese koopiaga tuberoossheroos II amplifitseerimisel.Selle keerulise malli puhul suurendas RNaseH-ravi SuperScript II või AMV-sünteesitud cDNA poolt toodetud signaali.Enamiku RT-PCR reaktsioonide puhul on RNaasH-ga töötlemine valikuline, kuna PCR-i denaturatsioonietapp 95 °C juures hüdrolüüsib üldiselt RNA-d RNA:DNA kompleksis.
6. Väikese RNA tuvastamise meetodi täiustamine:
RT-PCR on eriti keeruline, kui saadaval on ainult väikesed kogused RNA-d.RNA eraldamisel kandjana lisatud glükogeen aitab suurendada väikeste proovide saagist.RNaasivaba glükogeeni võib lisada samaaegselt Trizoli lisamisega.Glükogeen on vees lahustuv ja seda saab hoida vesifaasis koos RNA-ga, et soodustada järgnevat sadenemist.Proovide puhul, mis sisaldavad alla 50 mg koe või 106 kultiveeritud rakku, on RNaasivaba glükogeeni soovitatav kontsentratsioon 250 μg/ml.
Atsetüülitud BSA lisamine pöördtranskriptsioonireaktsioonile SuperScript II abil võib suurendada tundlikkust ning väikeste RNA koguste korral võib SuperScript II koguse vähendamine ja 40 ühiku RNaseOut nukleaasi inhibiitori lisamine tõsta tuvastamise taset.Kui RNA eraldamise protsessis kasutatakse glükogeeni, on SuperScript II kasutamisel pöördtranskriptsioonireaktsiooniks siiski soovitatav lisada BSA või RNaasi inhibiitor.
Näiteks RT-PCR spetsiifilisuse suurendamine
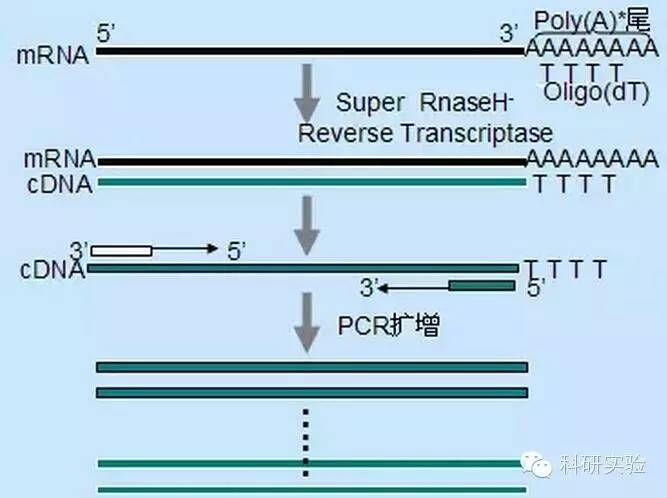
1. CND süntees:
Esimese ahela cDNA sünteesi saab algatada kolme erineva meetodi abil, mille suhteline spetsiifilisus mõjutab sünteesitava cDNA kogust ja tüüpi.
Juhusliku praimeri meetod oli kolmest meetodist kõige vähem spetsiifiline.Praimerid lõõmuvad kogu transkripti jooksul mitmes kohas, tekitades lühikesed, osalise pikkusega cDNA-d.Seda meetodit kasutatakse sageli 5'-otsa järjestuste saamiseks ja cDNA saamiseks RNA matriitsidest sekundaarse struktuuriga piirkondadega või terminatsioonisaitidega, mida ei saa pöördtranskriptaasi abil replitseerida.Pikima cDNA saamiseks tuleb empiiriliselt määrata praimerite ja RNA suhe igas RNA proovis.Juhuslike praimerite algkontsentratsioon oli vahemikus 50 kuni 250 ng 20 μl reaktsiooni kohta.Kuna juhuslike praimerite abil kogu RNA-st sünteesitud cDNA on peamiselt ribosomaalne RNA, valitakse matriitsiks üldiselt polü(A)+RNA.
Oligo(dT) praimerid on spetsiifilisemad kui juhuslikud praimerid.See hübridiseerub polü(A) sabaga, mida leidub enamiku eukarüootsete mRNAde 3'-otsas.Kuna polü(A)+ RNA on ligikaudu 1% kuni 2% kogu RNA-st, on cDNA kogus ja keerukus palju väiksem kui juhuslike praimerite puhul.Oma kõrge spetsiifilisuse tõttu ei nõua oligo(dT) üldiselt RNA ja praimerite suhte optimeerimist ja polü(A)+ selektsiooni.Soovitatav on kasutada 0,5 μg oligo(dT) 20 μl reaktsioonisüsteemi kohta.oligo(dT)12-18 sobib enamiku RT-PCR jaoks.ThermoScript RT-PCR süsteem pakub oligo(dT)20, kuna sellel on parem termiline stabiilsus kõrgematel inkubatsioonitemperatuuridel.
Geenispetsiifilised praimerid (GSP) on pöördtranskriptsiooni etapis kõige spetsiifilisemad praimerid.GSP on antisenss-oligonukleotiid, mis võib spetsiifiliselt hübridiseeruda RNA sihtjärjestusega, erinevalt juhuslikest praimeritest või oligo(dT-st), mis anniilivad kõigi RNA-dega.Samad reeglid, mida kasutatakse PCR praimerite kujundamisel, kehtivad GSP kavandamisel pöördtranskriptsioonireaktsioonides.GSP võib olla sama järjestus kui amplifikatsioonipraimer, mis anniilib mRNA kõige 3'-poolsema otsaga, või GSP võib olla konstrueeritud nii, et see anniiliks pöördamplifikatsioonipraimerist allavoolu.Mõnede amplifitseeritud subjektide jaoks tuleb edukaks RT-PCR-ks kavandada rohkem kui üks antisenss-praimer, kuna siht-RNA sekundaarne struktuur võib takistada praimeri seondumist.20 μl esimese ahela sünteesireaktsioonis on soovitatav kasutada 1 pmol antisenss GSP-d.
2. Tõstke pöördtranskriptsiooni inkubatsioonitemperatuuri:
GSP spetsiifilisuse täielikuks ärakasutamiseks tuleks kasutada suurema termostabiilsusega pöördtranskriptaasi.Termostabiilseid pöördtranskriptaase saab reaktsiooni ranguse suurendamiseks inkubeerida kõrgematel temperatuuridel.Näiteks kui GSP lõõmub temperatuuril 55 °C, ei kasutata GSP spetsiifilisust täielikult ära, kui AMV-d või M-MLV-d kasutatakse pöördtranskriptsiooniks madala rangusega 37 °C juures.SuperScript II ja ThermoScripti saab aga reageerida temperatuuril 50 °C või kõrgemal, mis kõrvaldab madalamatel temperatuuridel tekkivad mittespetsiifilised tooted.Maksimaalse spetsiifilisuse tagamiseks võib RNA/praimeri segu üle viia otse 65 °C denatureerimistemperatuurilt pöördtranskriptsiooni inkubatsioonitemperatuurile ja lisada eelsoojendatud 2x reaktsioonisegule (cDNA sünteesi kuumkäivitus).See aitab vältida molekulidevahelist aluste sidumist madalatel temperatuuridel.RT-PCR jaoks vajalikke mitut temperatuuri üleminekut saab lihtsustada termotsükleri abil.
3. Vähendab genoomse DNA saastumist:
RT-PCR-i võimalikuks raskuseks on genoomse DNA saastumine RNA-s.Hea RNA eraldamismeetodi (nt Trizol Reagent) kasutamine vähendab RNA preparaati saastava genoomse DNA kogust.Genoomsest DNA-st pärinevate toodete vältimiseks võib RNA-d töödelda amplifikatsiooniastmega DNaas I-ga, et eemaldada saastav DNA enne pöördtranskriptsiooni.DNaas I lõhustamine lõpetati, inkubeerides proove 2,0 mM EDTA-s 10 minutit temperatuuril 65 °C.EDTA võib kelaatida magneesiumioone, hoides ära magneesiumioonist sõltuva RNA hüdrolüüsi kõrgel temperatuuril.
Selleks, et eraldada amplifitseeritud cDNA saastavatest genoomse DNA amplifikatsiooniproduktidest, saab konstrueerida praimereid, mis kõik anniilivad eraldi eksonitega.cDNA-st pärinevad PCR-produktid on lühemad kui saastunud genoomsest DNA-st pärinevad.Lisaks viidi iga RNA matriitsiga läbi kontrollkatse ilma pöördtranskriptsioonita, et teha kindlaks, kas antud fragment pärineb genoomsest DNA-st või cDNA-st.Ilma pöördtranskriptsioonita saadud PCR-produkt pärineb genoomist.
Postitusaeg: mai-16-2023



